�о���ͬ������B-DNA��A-DNA����ת�����̲����Ľ��-985��ҵ����
��Դ��985������ ����ʱ�䣺2019-12-20 14:34
ժҪ�����÷��Ӷ���ѧģ�ⷽ���Ƚ������°�CHARMM��AMBER ������bsc1��OL15�� ������ˮ��Һ��B-DNA��A-DNAת�����̵�Ӱ�죬 ������չ����Ӧƫ���� ��eABF�� ����������ת�����̵������ܱ仯���о���������� �ڲ�ͬ�����£� ˮ�����е�DNA���ȶ��ṹ���ڲ��죬 AMBER������CHARMM����������ʵ������AMBER������DNA���ȶ��ṹ��С����խ�� �ȶ���B���ͣ���CHARMM������DNA���ȶ��ṹ��С���Ͽ��� ����B������A����֮�䡣ͨ������DNA��Χ���Ӽ�ˮ�ķֲ�������֣� CHARMM������DNAС����Χ�������ܶ����Ե���AMBER������ ���ܺܺõص���2������Ǽ�֮����ų����ã� ����CHARMM������С���Ͽ�������A���͵���Ҫԭ��
����
�����ؼ��ʣ�B-DNA; A-DNA; ����ת�䣻 CHARMM������ AMBER������ �����ܼ��㣻
����
����Abstract:The aim of the present work is to investigate and compare the effect of the latest CHARMM and AMBER force fields ��including bsc1 and OL15�� on the B-DNA to A-DNA conversion through exploring the free-energy changes of the conversion process. The extended adaptive biasing force ��e ABF�� method was utilized to perform the free-energy calculations. The results showed that the free-energy profiles characterizing the transition differ significantly for these two force fields. The AMBER force field performs better than the CHARMM force field in aqueous solution. The structure near the global minimum of the free-energy profile by the AMBER force field presents B-form, in agreement with the experimental results, while the most stable structure by the CHARMM force field locates between A-and B-form. Deep analysis of the radial distribution functions of the counterions around DNA reveals that the distribution of counterions in minor groove using the CHARMM force field is lower than that using the AMBER force field. Therefore, for the CHARMM force field, the repulsion of phosphates backbone could not be properly counteracted by counterions, as a result, the minor groove becomes wider, causing a slight conformational change towards A-form.
����
����Keyword:B-DNA; A-DNA; Conformational conversion; CHARMM force field; AMBER force field; Free-energy calculation;
 ����
����
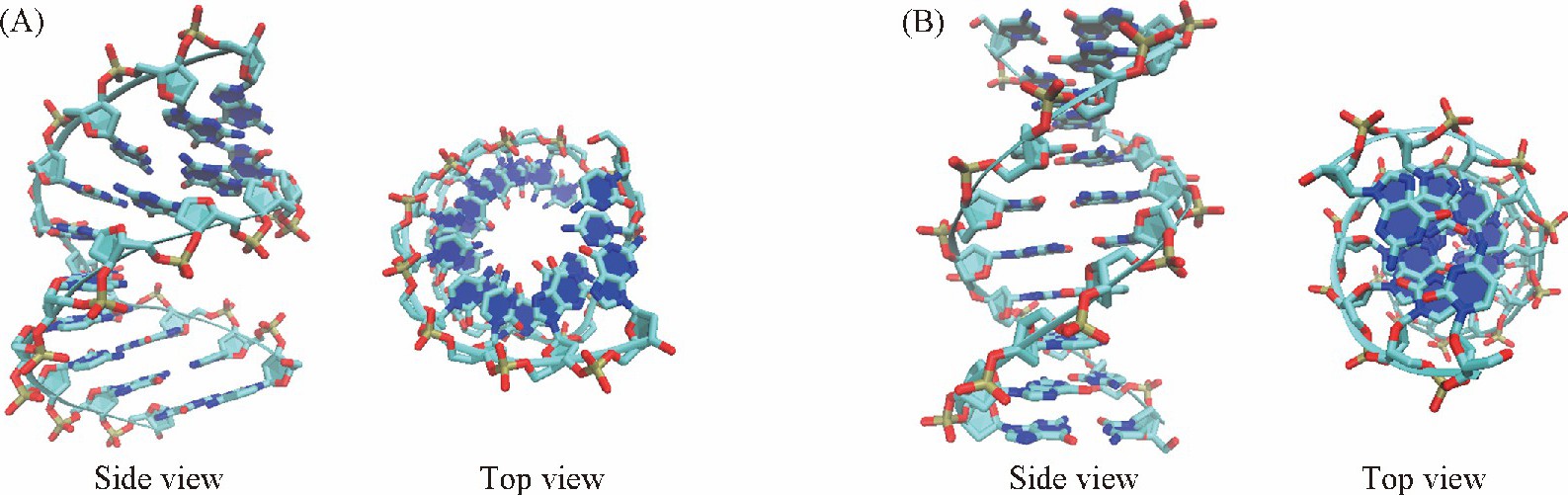
����DNA��Ϊ������Ϣ���ݵ����壬 ��Ҫ����B-DNA, A-DNA��Z-DNA 3����ʽ[1].ͨ���� ��ˮ��ȽϸߵĻ��� ����ϸ���� �У� DNA��Ҫ��B���͵���ʽ���ڡ�A-DNA����Ҫ������ˮ��Ƚϵ͵Ļ��� ������λ���Ҵ���Һ�� �У� ��DNA��RNA���ʱ�� ͬ�������A����[2].B������A����DNA�Ľṹ��ͼ1��ʾ�� ���߾���������״̬�� ��A����С���Ͽ��� �����϶̣� ���ָ����ܵ�����״̬��DNA�ṹ�ĸı��������ϢϢ��أ� ���Ӷ���ѧ ��MD�� ģ�ⱻ�㷺Ӧ�����о�DNA�ṹ�ı�Ļ���[3,4].�����Ӷ���ѧģ���ȷ�Լ���̶��������ڷ��������ľ�ȷ�̶ȡ�CHARMM[5,6], AMBER[7,8], GROMOS[9]��OPLS[10]������Ӧ��������������ϵ�ѽ�Ϊ���졣����DNA, Ӧ�ýϹ㷺����������CHARMM��AMBER������ ����AMBER�����е�bsc1[7]��OL15[8]������Ϊ����DNA�����°��������㷺Ӧ�á����������IJ����Ż��뷢չ�� �������ȷ�Բ�����ߣ� ��������Ȼ���ڣ� ��˷��ֲ��Ľ����������IJ������Ż�����������Ŀ�ꡣ�����о���������� CHARMM��������DNA, ��ṹ������A���ͣ� AMBER���ȶ���B����[11].��ˣ� ��ͬ������Ӧ�÷�Χ���ڲ��죬 ͨ���� ��ˮ������AMBER�����ܹ����õ�����B���ͣ� ����ˮ��Ƚϵ͵Ļ����У� CHARMM������A-DNA�����������ȷ�� ��ʵ�������Ǻ�[12,13].��Ȼ��ͬ�����µĽṹ��Ϣ�о��ñȽ���ϸ�� ���Ը�����ı��ʽ��Ͳ���ʮ�������ͬʱ�� �������������ķ�չ�� �������µ�������������㷺Ӧ��[7,8].
 ����
����
����Ϊ�������о����°�CHARMM��AMBER������DNA����ת���Ӱ�죬 ���IJ��÷���ģ���������ܼ���ķ���[14,15]�Ƚ��˲�ͬ������ˮ��Һ��B-DNA��A-DNAת���������ܱ仯��ͨ�������ܼ������ʵ�ֶ�B���͵�A����ת����̵ķ����� ��������� CHARMM�����µ�DNAС���Ͽ��� ��ʵ�������ڲ��죬 ���þ���ֲ������Ը�������з����� ���ֲ�ͬ������DNA��Χ�����ӷֲ����ڽϴ����� ��ͬ�����ӷֲ������DNA�ȶ��ṹ���ڲ������Ҫ���ء�
����
����1�����ۺͷ���
����
����1.1 ģ�ͽ���
����
����B-DNA��A-DNA�ij�ʼ�ṹ��AMBER�ṩ��Nucleic Acid Builder��������� ���зֱ�ΪGCGCGC��ATATAT.�������õ�DNAģ�����ڴ��������Ա߽�������ˮ���������У� ÿ����ϵ����10��K+��Ϊ�������ӣ� ���Ӵ�СԼΪ6.1 nm×6.0 nm×6.4 nm, ÿ����ϵ��ԭ������Ϊ20000�����ҡ�����ģ����Ȼ������������AT���� ��PDB code:4J2I�� �Լ�GC���� ��PDB code:1QC1�� �ľ���ṹ�ο��Ե����ʾ���ṹ���ݿ�[16,17].
����
����1.2 ����ѧģ��
����
��������NAMD2.12��������MDģ��[18], �ֱ�ʹ��CHARMM36������AMBER-OL15������AMBER-bsc1������������DNA;ʹ��TIP3Pģ��[19]�еIJ�������ˮ���ӡ�����SKAKE/RATTLE�㷨[20,21]����ˮ�����к�����ԭ�ӵĹ��ۼ��ij�����������ƽ��ֵ�� ����SETTLE�㷨[22]����ˮ���ӵĸ��ԡ����ú��º�ѹ��Langevin����ѧ������Langevin��������[23]���¶Ⱥ�ѹ���ֱ������300 K��1.01×105Pa.���»��ضϰ뾶Ϊ1.2 nm, ���̾�������ò������������߶��� ��PME�� �������㡣���˶����̻��ֵ�ʱ�䲽��Ϊ2 fs, ÿ����ϵ�������ܼ���֮ǰ��������2000����������С�����̡��������ܼ�������ѡȡ���ȶ��ṹ����10 ns��ƽ��ģ�⣬ Ӧ�øù켣����DNA�ṹ�IJ����������������Ӻ�ˮ���ӵľ���ֲ�����������VMD����[24]���й켣������Ӧ��CURVES+����[25]����DNA�ṹ���������� ����������DNA��ĩ�˷ֱ��ȥ���Լ���� ѡȡ�м�л����з�����
����
����1.3 �����ܼ���
����
����������չ����Ӧƫ���� ��e ABF�� ����������ת�����̵������ܱ仯[15], ��ΔRMSD ��RMSDB-RMSDA�� ��Ϊ��ת�����̵ķ�Ӧ����[26].ѡȡDNA��������ԭ����ΪRMSD����IJο�ԭ�ӣ� ��B-DNA���A-DNA��RMSD��ֵΪ0.3 nm, -0.3<ΔRMSD<0.3.��ΔRMSD<0ʱ�� DNA�Ľṹƫ����B���ͣ���ΔRMSD>0ʱ�� DNA�ṹƫ����A���͡�Ϊ�������Ч�ʣ� ��Ӧ·��������Ϊ2�����ڣ�-0.3<ΔRMSD<0��0<ΔRMSD<0.3.ÿ�����ڵ�ģ��ʱ��Ϊ150 ns, ÿ����ϵ��ģ��ʱ��Ϊ300 ns, ��ģ��ʱ��Ϊ1.2μs.
����
����2�����������
����
����2.1 ����������
����
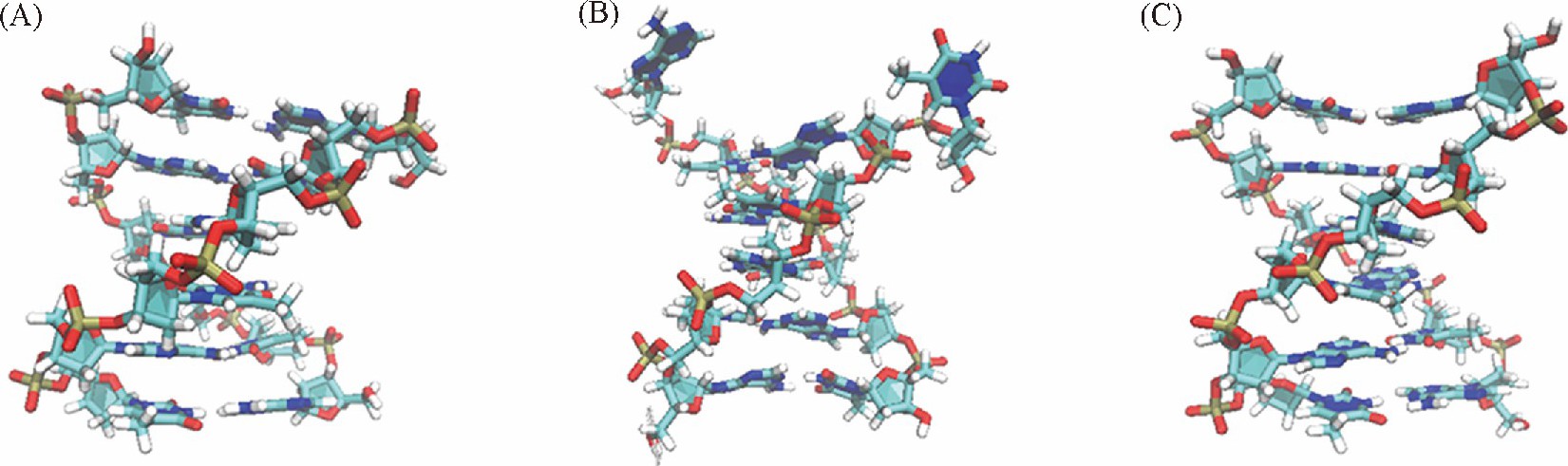
����ͼ2 ��A�� ������ATATAT����B-DNA��A-DNAת��������ܱ仯������AMBRER���� ������bsc1������OL15������ , ���ȶ��ĽṹΪB-DNA, bsc1������OL15������Ӧ��ΔRMSDֵ�ֱ�Ϊ-0.14��-0.15 nm, ��ͼ3 ��A�� �� ��C�� ��ʾ�� ��ṹ�dz����ơ�����CHARMM������ ����һ����Χ�Ϲ�ĵ������� ����ȡ������͵�Ľṹ[ͼ3 ��B�� ]�ɼ��� DNA��ĩ�˵�����ƻ���Ϊ���أ� ������ƻ�ʹĩ�˼���˶���Ϊ�� ���ǵ������ȶ��ṹΔRMSDֵ��Χ�Ϲ��ԭ��֮һ��
����
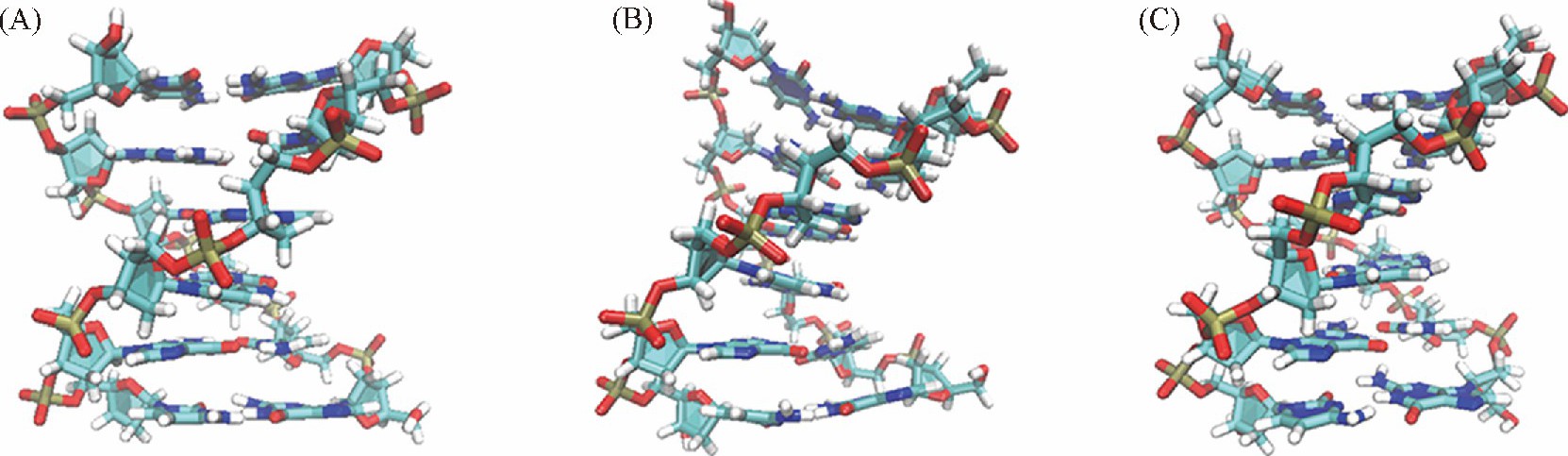
����ͼ2 ��B�� ������GCGCGC���е�B-DNA��A-DNAת��������ܱ仯���ɼ��� ��ˮ�����в�ͬ������DNA�����ȶ��ṹ���ڲ��졣��ͼ4 ��A�� �� ��C�� ��ʾ�� ����AMBER����������bsc1��������OL15������ B-DNA��A-DNAת���Ĺ��������ȶ��ĽṹΪB-DNA, ��Ӧ��ΔRMSDֵ�ֱ�Ϊ-0.12��-0.13 nm.����CHARMM������ ���ȶ��Ľṹ��ƫ�����B-DNA, ����Ϊ����B������A�����м�Ľṹ[ͼ4 ��B�� ], ��Ӧ��ΔRMSDֵΪ-0.02 nm, ��ʵ���ϱ��ֳ����ı�B-DNA���ڲ��졣���Ͻ����ʾ�� AMBER����������ˮ��Һ��B-DNA��A-DNAת�������ȶ��ṹ����ȷ�� �ý����ɰ�������ģ��������[12,13], ��bsc1������OL15�������������ͬ�� ��ˣ� ����ֻѡȡbsc1�������з�����
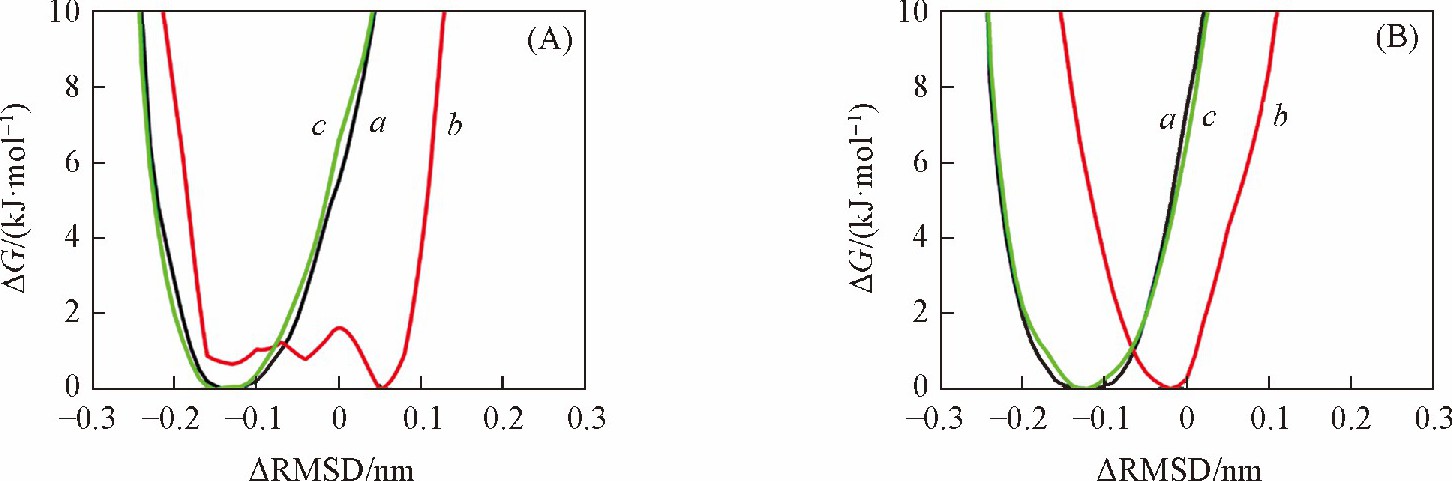
 ����
����
����2.2 DNA�ṹ��������
����
����Ϊ���о��ڲ�ͬ������DNAȫ��������С����ȶ��ṹ�� ��ȡ��ȫ��������͵�Ĵ����ṹ����ƽ��ģ�Ⲣ������ṹ����������ͬ����DNA�Ľṹ�����뾧��ṹ�����˱Ƚ� ����S1, ������֧����Ϣ�� , ��������� ���� ��AT�� 6���У� AMBER�����µľ��ֽṹ�����ܹ����ӽ�ʵ������������ ��GC�� 6���У� CHARMM��AMBER�����Բ�ͬ�����ľ�ȷ�̶Ȳ�ͬ�����в�ͬ�������ϴ���Ǵ�С���Ŀ��ȣ� Ϊ�˽�һ��̽����ԭ�� ���������������
����
����Ϊ�˽�һ��̽��DNA�ڲ�ͬ�����µĽṹ���죬 ��DNA��С���Ŀ��Ƚ�����ͳ�Ʒ�����ͼ5�����˲�ͬ�����²�ͬ������еĴ��ȷֲ������ ���ֶ��� ��AT�� 6���У� bsc1�����µĴ��ȷֲ���ΧΪ1.1~1.4 nm, �뾧��ṹ�����Ǻϣ���CHARMM��������ֽ�խ�Ĵ��ȣ� �뾧��ṹ���ڲ���[ͼ5 ��A�� ].���� ��GC�� 6���У� bsc1�����µĴ�������ھ���ṹ�Կ��� CHARMM�����µĴ����뾧��ṹ��Ϊ����[ͼ5 ��B�� ].
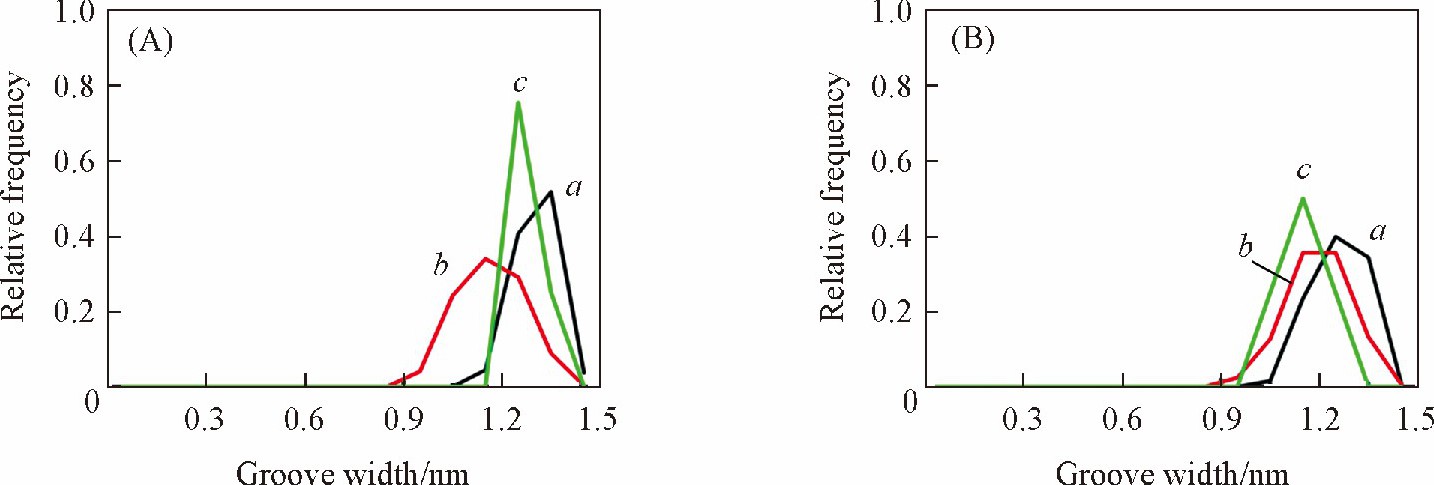 ����
����
����ͼ6������DNAС�����ȵķֲ������ ��AT�� 6���о���ṹ��С�����ȷֲ���Χ��0.3~0.6 nm֮�䣬 bsc1������һ���̶����ܹ���ӳ����ṹ��С�����ȣ� ������Ȼ����С������ƫ�����������CHARMM������ С�����ȷֲ���0.6~1.3 nm֮�䣬 �ý����ȫƫ�뾧��ṹ�ṩ����Ϣ[ͼ6 ��A�� ].���� ��GC�� 6���У� bsc1�����ܴ�һ���̶��Ϸ�ӳ����ṹ��С�����ȣ� ��CHARMM�����µ�С�������ڷֲ����������ƣ� ���ֽϿ���С������[ͼ6 ��B�� ].�ɼ��� bsc1������DNAС�����ȵ�������Ϊȷ�� ��CHARMM�����²�ͬ���е�С�����Ⱦ����ھ���ṹ���ɴ˿ɼ��� AMBER���� ��bsc1�� ������DNA��С���Ŀ������Ա�CHARMM�������Ӿ�ȷ��
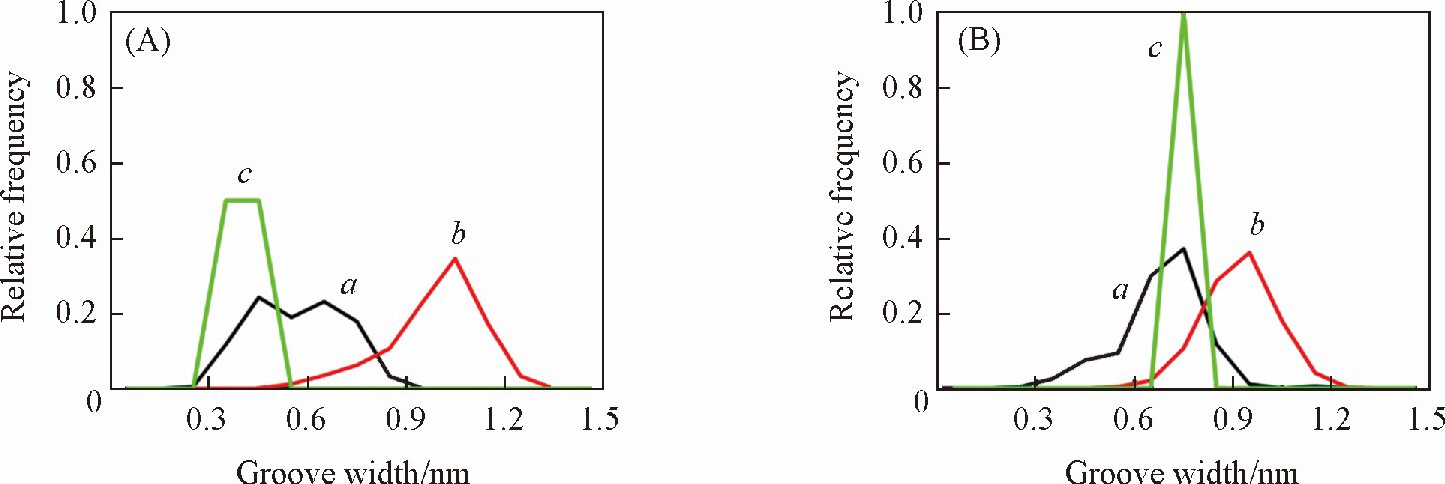 ����
����
����2.3 �������Ӷ�DNA�ṹ��Ӱ��
����
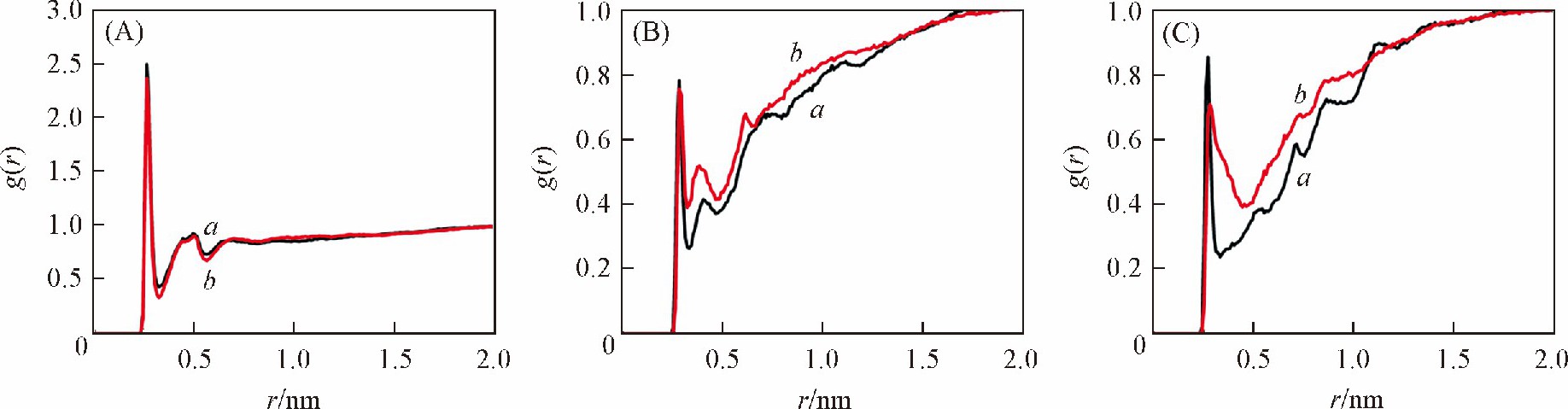
����Ϊ��̽����ͬ�����´�С�����ȴ��ڽϴ�����ԭ�� ��DNA��Χ�������ӵķֲ�������̽����ͼ7��ͼ8�ֱ������� ��GC�� 6������ ��AT�� 6�����ڲ�ͬ������DNA�Ǽܡ���С����Χ�����ӷֲ��������ͼ7 ��A�� , ��C�� ��ͼ8 ��A�� , ��C�� �ɼ��� ������ ��GC�� 6���л��� ��AT�� 6���У� bsc1�����¹Ǽܼ�С����Χ�����ӷֲ��ܶȾ����Ը���CHARMM������bsc1�����½��ܵ����ӷֲ����õ��к�������Ǽ��ϵĸ���ɣ� ������2������Ǽ�֮��ľ����ų����ã� �����CHARMM������ȣ� bsc1�����µ�DNA�ṹС���Ŀ��Ƚ�խ���ɼ��� CHARMM��������ȷ����С�����ȵ���Ҫ�����ǣ� ��CHARMM��������������С���ķֲ����٣� ���´�����ɵ�����Ǽܾ����ų�ϴ� С�������
����
������ͼ7 ��B�� ��ͼ8 ��B�� �ɼ��� ������ͬ���У� bsc1������CHARMM������������DNA����Χ�ķֲ��ܶȲ�� Ȼ�� ��GC�� 6������ ��AT�� 6����֮��ȴ���ڽϴ���졣��������[27]������ ���� ��GC�� 6���е�DNA, �����ڴķֲ��ܶȸ��� ��AT�� 6���У� ����ģ����Ҳ�õ��˺ܺõ���֤��ͬʱ�� С����Χ�����ӷֲ����ڲ�ͬ�ļ������Ҳ���ϴ� ��ͼ7 ��C�� ��ͼ8 ��C�� �ɼ��� ���� ��AT�� 6���е�DNA, ������С���ֲ����������Ը��� ��GC�� 6���У� ������С����Χ�Ľ��ֲܷ����õ��к�������Ǽܵĸ���ɣ� �����˾����ų����ã� ʹС����խ�� ��ܺõؽ�����ʵ���� ��AT�� 6����DNA��С������С�� ��GC�� 6���е����ɼ��� ������DNA��Χ�ķֲ������Ӱ��DNA��С�����ȵ���Ҫ���ء�
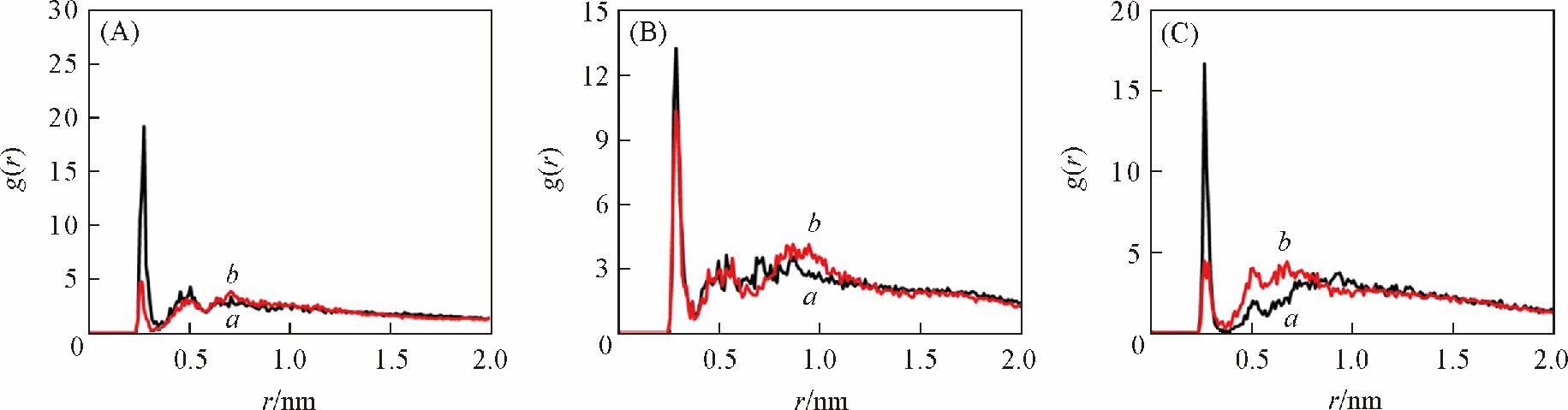
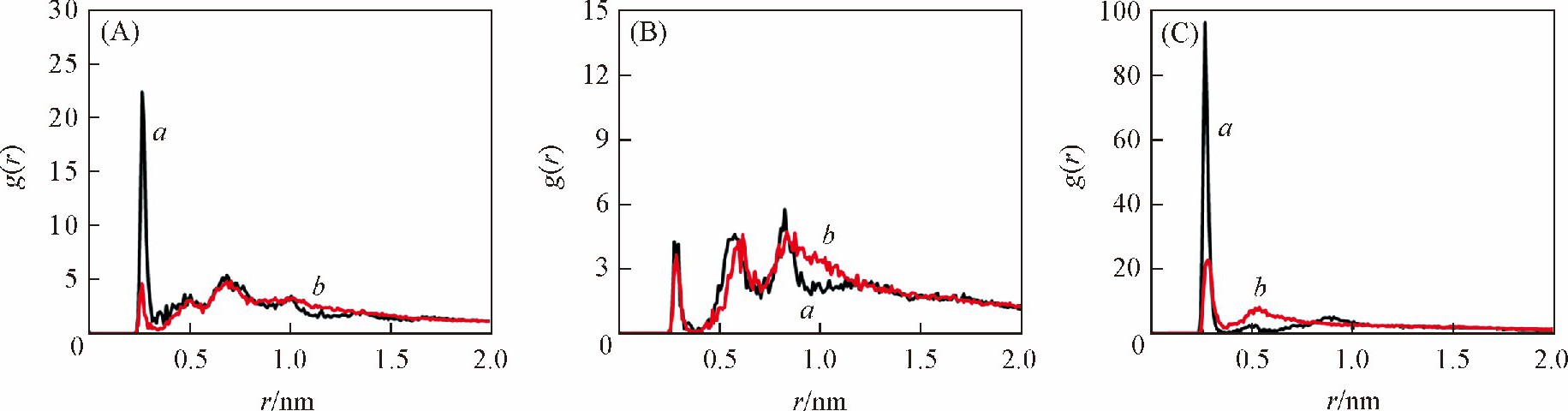 ����
����
����2.4ˮ���Ӷ�DNA�ṹ��Ӱ��
����
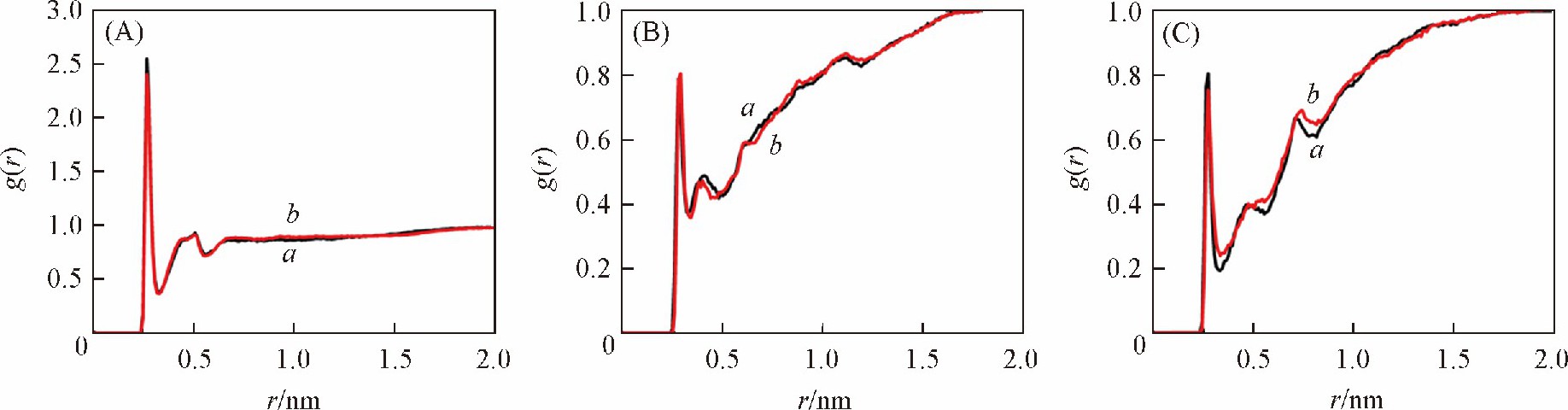
����DNA��Χˮ���ӵķֲ����ͬ����DNA�Ľṹ���нϴ��Ӱ�졣���� ��GC�� 6���У� ��ͬ������DNA��Χˮ���ӵķֲ����� ��ͼ9�� ;������ ��AT�� 6���У� CHARMM������DNA�Ĵ�С����Χ��ˮ���ӷֲ��϶� ��ͼ10�� .Ϊ��̽��ˮ���Ӷ�DNA�ṹ��Ӱ�죬 ������ȡ��ƽ��ģ����DNA�ĵ��ͽṹ�� ��ͼ3 ��B�� �ɼ��� DNAĩ�˼�������ˮ�����ƻ���Ϊ���ء���ˣ� ����CHARMM������ ��AT�� 6����ĩ�˼��������ױ���Χ��ˮ�����ƻ��� ��Ҳ�� ��AT�� 6���д�С����Χˮ���ӷֲ��϶��ԭ���ڼ��֮�����3������� ��GC�� 6���У� ������γ�2������� ��AT�� 6���и����ȶ��� CHARMM������bsc1������DNA��Χˮ���ӵķֲ������û�нϴ�����
 ����
����
����3������
����
�������÷��Ӷ���ѧģ���������ܼ���ķ����Ƚ���CHARMM��AMBER���� ������bsc1������OL15������ ��ˮ��Һ��B-DNA��A-DNAת�����̵�Ӱ�죬 �ֱ�õ��˲�ͬ���������ȶ���DNA�ṹ����������� ��ˮ�����в�ͬ������DNA�����ȶ��ṹ���ڲ��죬 CHARMM������DNA���ȶ��ṹ��С���Ͽ��� ����B������A����֮�䣻AMBER�����µ�DNAС����խ�� �ȶ���B���͡���һ��̽���˲�ͬ������DNA�ȶ��ṹ���ڲ����ԭ�� ���ȷ�����DNA��Χ���ӵķֲ��������������� CHARMM������DNAС����Χ�������ܶ����Ե���AMBER������ ���ܺܺõص�������Ǽܵ��ų����ã� ��CHARMM����С���Ͽ�������A���͵���Ҫԭ��һ��������ˮ���ӵķֲ���DNA�ṹ��Ӱ�죬 ����CHARMM�����µ� ��AT�� 6����ĩ�˼��֮��������ˮ�����ƻ���Ϊ���أ� ��AMBER������ȣ� CHARMM�����µ��������ý�������ģ������������ˮ��Һ��AMBER�����ܹ����õ�����ʵ�������о����Ϊ����ģ��ʵ������ṩ������ָ���� ���Խ�һ���IJ����Ż��ṩ�˰�����
����
���������
����[1]Saenger W., Defining Terms for the Nucleic Acids, Springer, New York, 1984
����[2]Ivanov V.I., Minchenkova L.E., Minyat E.E., Frank-Kamenetskii M.D., Schyolkina A.K., J.Mol.Biol., 1974, 87 ��4�� , 817-833
����[3]Mukherjee A., Lavery R., Bagchi B., Hynes J.T., J.Am.Chem.Soc., 2008, 130 ��30�� , 9747-9755
����[4]Dumont E., Wibowo M., Roca-Sanjun D., Garavelli M., Assfeld X., Monari A., J.Phys.Chem.Lett., 2015, 6 ��4�� , 576-580
����[5]Foloppe N., Mac Kerell A.D.Jr., J.Comput.Chem., 2000, 21 ��2�� , 86-104
����[6]Mac Kerell A.D.Jr., Bashford D., Bellott M., Dunbrack R.L.Jr., Evanseck J.D., Field M.J., Fischer S., Gao J., Guo H., Ha S., J.Phys.Chem.B, 1998, 102 ��18�� , 3586-3616
����[7]Ivani I., Dans P.D., Noy A., Pérez A., Faustino I., Hospital A., Walther J., Andrio P., Go1i R., Balaceanu A., Nat.Methods, 2016, 13 ��1�� , 55
����[8]ZgarbováM., Luque F.J., poner J.I., Cheatham III T.E., Otyepka M., Jurecka P., J.Chem.Theory Comput., 2013, 9 ��5�� , 2339-2354
����[9]Soares T.A., Hünenberger P.H., Kastenholz M.A., Krutler V., Lenz T., Lins R.D., Oostenbrink C., van Gunsteren W.F., J.Comput.Chem., 2005, 26 ��7�� , 725-737
����[10]Robertson M.J., Tirado-Rives J., Jorgensen W.L., J.Chem.Theory Comput., 2015, 11 ��7�� , 3499-3509
����[11]Song C., Xia Y.Y., Zhao M.W., Liu X.D., Li F., Ji Y.J., Huang B.D., Yin Y.Y., J.Mol.Model., 2006, 12 ��3�� , 249-254
����[12]Cheatham T.E., Crowley M.F., Fox T., Kollman P.A., Proc.Natl.Acad.Sci.USA, 1997, 94 ��18�� , 9626-9630
����[13]Yang L., Pettitt B.M., J.Phys.Chem., 1996, 100 ��7�� , 2564-2566
����[14]Shen W., Shao X.G., Cai W.S., Chem.J.Chinese Universities, 2016, 37 ��10�� , 1809-1816 �����ģ� ��ѧ�㣬 ���������ߵ�ѧУ��ѧѧ���� 2016, 37 ��10�� , 1809-1816��
����[15]Fu H.H., Shao X.G., Chipot C., Cai W.S., J.Chem.Theory Comput., 2016, 12 ��8�� , 3506-3513
����[16]Acosta-Reyes F.J., Subirana J.A., Pous J., Sánchez-Giraldo R., Condom N., Baldini R., Malinina L., Campos J.L., Biopolymers, 2015, 103 ��3�� , 123-133
����[17]Timsit Y., Moras D., EMBO J., 1994, 13 ��12�� , 2737
����[18]Phillips J.C., Braun R., Wang W., Gumbart J., Tajkhorshid E., Villa E., Chipot C., Skeel R.D., Kale L., Schulten K., J.Comput.Chem., 2005, 26 ��16�� , 1781-1802
����[19]Jorgensen W.L., Chandrasekhar J., Madura J.D., Impey R.W., Klein M.L., J.Chem.Phys., 1983, 79 ��2�� , 926-935
����[20]Ryckaert J.P., Ciccotti G., Berendsen H.J., J.Comput.Phys., 1977, 23 ��3�� , 327-341
����[21]Andersen H.C., J.Comput.Phys., 1983, 52 ��1�� , 24-34
����[22]Miyamoto S., Kollman P.A., J.Comput.Chem., 1992, 13 ��8�� , 952-962
����[23]Feller S.E., Zhang Y., Pastor R.W., Brooks B.R., J.Chem.Phys., 1995, 103 ��11�� , 4613-4621
����[24]Humphrey W., Dalke A., Schulten K., J.Mol.Graphics, 1996, 14 ��1�� , 33-38
����[25]Lavery R., Moakher M., Maddocks J., Petkeviciute D., Zakrzewska K., Nucleic Acids Res., 2009, 37 ��17�� , 5917-5929
����[26]Banavali N.K., Roux B., J.Am.Chem.Soc., 2005, 127 ��18�� , 6866-6876
����[27]Hamelberg D., Williams L.D., Wilson W.D., J.Am.Chem.Soc., 2001, 123 ��32�� , 7745-7755
����
�����ؼ��ʣ�B-DNA; A-DNA; ����ת�䣻 CHARMM������ AMBER������ �����ܼ��㣻
����
����Effect of Different Force Fields on B-DNA to A-DNA Conversion�k
��������Abstract:The aim of the present work is to investigate and compare the effect of the latest CHARMM and AMBER force fields ��including bsc1 and OL15�� on the B-DNA to A-DNA conversion through exploring the free-energy changes of the conversion process. The extended adaptive biasing force ��e ABF�� method was utilized to perform the free-energy calculations. The results showed that the free-energy profiles characterizing the transition differ significantly for these two force fields. The AMBER force field performs better than the CHARMM force field in aqueous solution. The structure near the global minimum of the free-energy profile by the AMBER force field presents B-form, in agreement with the experimental results, while the most stable structure by the CHARMM force field locates between A-and B-form. Deep analysis of the radial distribution functions of the counterions around DNA reveals that the distribution of counterions in minor groove using the CHARMM force field is lower than that using the AMBER force field. Therefore, for the CHARMM force field, the repulsion of phosphates backbone could not be properly counteracted by counterions, as a result, the minor groove becomes wider, causing a slight conformational change towards A-form.
����
����Keyword:B-DNA; A-DNA; Conformational conversion; CHARMM force field; AMBER force field; Free-energy calculation;
����DNA��Ϊ������Ϣ���ݵ����壬 ��Ҫ����B-DNA, A-DNA��Z-DNA 3����ʽ[1].ͨ���� ��ˮ��ȽϸߵĻ��� ����ϸ���� �У� DNA��Ҫ��B���͵���ʽ���ڡ�A-DNA����Ҫ������ˮ��Ƚϵ͵Ļ��� ������λ���Ҵ���Һ�� �У� ��DNA��RNA���ʱ�� ͬ�������A����[2].B������A����DNA�Ľṹ��ͼ1��ʾ�� ���߾���������״̬�� ��A����С���Ͽ��� �����϶̣� ���ָ����ܵ�����״̬��DNA�ṹ�ĸı��������ϢϢ��أ� ���Ӷ���ѧ ��MD�� ģ�ⱻ�㷺Ӧ�����о�DNA�ṹ�ı�Ļ���[3,4].�����Ӷ���ѧģ���ȷ�Լ���̶��������ڷ��������ľ�ȷ�̶ȡ�CHARMM[5,6], AMBER[7,8], GROMOS[9]��OPLS[10]������Ӧ��������������ϵ�ѽ�Ϊ���졣����DNA, Ӧ�ýϹ㷺����������CHARMM��AMBER������ ����AMBER�����е�bsc1[7]��OL15[8]������Ϊ����DNA�����°��������㷺Ӧ�á����������IJ����Ż��뷢չ�� �������ȷ�Բ�����ߣ� ��������Ȼ���ڣ� ��˷��ֲ��Ľ����������IJ������Ż�����������Ŀ�ꡣ�����о���������� CHARMM��������DNA, ��ṹ������A���ͣ� AMBER���ȶ���B����[11].��ˣ� ��ͬ������Ӧ�÷�Χ���ڲ��죬 ͨ���� ��ˮ������AMBER�����ܹ����õ�����B���ͣ� ����ˮ��Ƚϵ͵Ļ����У� CHARMM������A-DNA�����������ȷ�� ��ʵ�������Ǻ�[12,13].��Ȼ��ͬ�����µĽṹ��Ϣ�о��ñȽ���ϸ�� ���Ը�����ı��ʽ��Ͳ���ʮ�������ͬʱ�� �������������ķ�չ�� �������µ�������������㷺Ӧ��[7,8].
����Ϊ�������о����°�CHARMM��AMBER������DNA����ת���Ӱ�죬 ���IJ��÷���ģ���������ܼ���ķ���[14,15]�Ƚ��˲�ͬ������ˮ��Һ��B-DNA��A-DNAת���������ܱ仯��ͨ�������ܼ������ʵ�ֶ�B���͵�A����ת����̵ķ����� ��������� CHARMM�����µ�DNAС���Ͽ��� ��ʵ�������ڲ��죬 ���þ���ֲ������Ը�������з����� ���ֲ�ͬ������DNA��Χ�����ӷֲ����ڽϴ����� ��ͬ�����ӷֲ������DNA�ȶ��ṹ���ڲ������Ҫ���ء�
����
����1�����ۺͷ���
����
����1.1 ģ�ͽ���
����
����B-DNA��A-DNA�ij�ʼ�ṹ��AMBER�ṩ��Nucleic Acid Builder��������� ���зֱ�ΪGCGCGC��ATATAT.�������õ�DNAģ�����ڴ��������Ա߽�������ˮ���������У� ÿ����ϵ����10��K+��Ϊ�������ӣ� ���Ӵ�СԼΪ6.1 nm×6.0 nm×6.4 nm, ÿ����ϵ��ԭ������Ϊ20000�����ҡ�����ģ����Ȼ������������AT���� ��PDB code:4J2I�� �Լ�GC���� ��PDB code:1QC1�� �ľ���ṹ�ο��Ե����ʾ���ṹ���ݿ�[16,17].
����
����1.2 ����ѧģ��
����
��������NAMD2.12��������MDģ��[18], �ֱ�ʹ��CHARMM36������AMBER-OL15������AMBER-bsc1������������DNA;ʹ��TIP3Pģ��[19]�еIJ�������ˮ���ӡ�����SKAKE/RATTLE�㷨[20,21]����ˮ�����к�����ԭ�ӵĹ��ۼ��ij�����������ƽ��ֵ�� ����SETTLE�㷨[22]����ˮ���ӵĸ��ԡ����ú��º�ѹ��Langevin����ѧ������Langevin��������[23]���¶Ⱥ�ѹ���ֱ������300 K��1.01×105Pa.���»��ضϰ뾶Ϊ1.2 nm, ���̾�������ò������������߶��� ��PME�� �������㡣���˶����̻��ֵ�ʱ�䲽��Ϊ2 fs, ÿ����ϵ�������ܼ���֮ǰ��������2000����������С�����̡��������ܼ�������ѡȡ���ȶ��ṹ����10 ns��ƽ��ģ�⣬ Ӧ�øù켣����DNA�ṹ�IJ����������������Ӻ�ˮ���ӵľ���ֲ�����������VMD����[24]���й켣������Ӧ��CURVES+����[25]����DNA�ṹ���������� ����������DNA��ĩ�˷ֱ��ȥ���Լ���� ѡȡ�м�л����з�����
����
����1.3 �����ܼ���
����
����������չ����Ӧƫ���� ��e ABF�� ����������ת�����̵������ܱ仯[15], ��ΔRMSD ��RMSDB-RMSDA�� ��Ϊ��ת�����̵ķ�Ӧ����[26].ѡȡDNA��������ԭ����ΪRMSD����IJο�ԭ�ӣ� ��B-DNA���A-DNA��RMSD��ֵΪ0.3 nm, -0.3<ΔRMSD<0.3.��ΔRMSD<0ʱ�� DNA�Ľṹƫ����B���ͣ���ΔRMSD>0ʱ�� DNA�ṹƫ����A���͡�Ϊ�������Ч�ʣ� ��Ӧ·��������Ϊ2�����ڣ�-0.3<ΔRMSD<0��0<ΔRMSD<0.3.ÿ�����ڵ�ģ��ʱ��Ϊ150 ns, ÿ����ϵ��ģ��ʱ��Ϊ300 ns, ��ģ��ʱ��Ϊ1.2μs.
����
����2�����������
����
����2.1 ����������
����
����ͼ2 ��A�� ������ATATAT����B-DNA��A-DNAת��������ܱ仯������AMBRER���� ������bsc1������OL15������ , ���ȶ��ĽṹΪB-DNA, bsc1������OL15������Ӧ��ΔRMSDֵ�ֱ�Ϊ-0.14��-0.15 nm, ��ͼ3 ��A�� �� ��C�� ��ʾ�� ��ṹ�dz����ơ�����CHARMM������ ����һ����Χ�Ϲ�ĵ������� ����ȡ������͵�Ľṹ[ͼ3 ��B�� ]�ɼ��� DNA��ĩ�˵�����ƻ���Ϊ���أ� ������ƻ�ʹĩ�˼���˶���Ϊ�� ���ǵ������ȶ��ṹΔRMSDֵ��Χ�Ϲ��ԭ��֮һ��
����
����ͼ2 ��B�� ������GCGCGC���е�B-DNA��A-DNAת��������ܱ仯���ɼ��� ��ˮ�����в�ͬ������DNA�����ȶ��ṹ���ڲ��졣��ͼ4 ��A�� �� ��C�� ��ʾ�� ����AMBER����������bsc1��������OL15������ B-DNA��A-DNAת���Ĺ��������ȶ��ĽṹΪB-DNA, ��Ӧ��ΔRMSDֵ�ֱ�Ϊ-0.12��-0.13 nm.����CHARMM������ ���ȶ��Ľṹ��ƫ�����B-DNA, ����Ϊ����B������A�����м�Ľṹ[ͼ4 ��B�� ], ��Ӧ��ΔRMSDֵΪ-0.02 nm, ��ʵ���ϱ��ֳ����ı�B-DNA���ڲ��졣���Ͻ����ʾ�� AMBER����������ˮ��Һ��B-DNA��A-DNAת�������ȶ��ṹ����ȷ�� �ý����ɰ�������ģ��������[12,13], ��bsc1������OL15�������������ͬ�� ��ˣ� ����ֻѡȡbsc1�������з�����
����2.2 DNA�ṹ��������
����
����Ϊ���о��ڲ�ͬ������DNAȫ��������С����ȶ��ṹ�� ��ȡ��ȫ��������͵�Ĵ����ṹ����ƽ��ģ�Ⲣ������ṹ����������ͬ����DNA�Ľṹ�����뾧��ṹ�����˱Ƚ� ����S1, ������֧����Ϣ�� , ��������� ���� ��AT�� 6���У� AMBER�����µľ��ֽṹ�����ܹ����ӽ�ʵ������������ ��GC�� 6���У� CHARMM��AMBER�����Բ�ͬ�����ľ�ȷ�̶Ȳ�ͬ�����в�ͬ�������ϴ���Ǵ�С���Ŀ��ȣ� Ϊ�˽�һ��̽����ԭ�� ���������������
����
����Ϊ�˽�һ��̽��DNA�ڲ�ͬ�����µĽṹ���죬 ��DNA��С���Ŀ��Ƚ�����ͳ�Ʒ�����ͼ5�����˲�ͬ�����²�ͬ������еĴ��ȷֲ������ ���ֶ��� ��AT�� 6���У� bsc1�����µĴ��ȷֲ���ΧΪ1.1~1.4 nm, �뾧��ṹ�����Ǻϣ���CHARMM��������ֽ�խ�Ĵ��ȣ� �뾧��ṹ���ڲ���[ͼ5 ��A�� ].���� ��GC�� 6���У� bsc1�����µĴ�������ھ���ṹ�Կ��� CHARMM�����µĴ����뾧��ṹ��Ϊ����[ͼ5 ��B�� ].
����ͼ6������DNAС�����ȵķֲ������ ��AT�� 6���о���ṹ��С�����ȷֲ���Χ��0.3~0.6 nm֮�䣬 bsc1������һ���̶����ܹ���ӳ����ṹ��С�����ȣ� ������Ȼ����С������ƫ�����������CHARMM������ С�����ȷֲ���0.6~1.3 nm֮�䣬 �ý����ȫƫ�뾧��ṹ�ṩ����Ϣ[ͼ6 ��A�� ].���� ��GC�� 6���У� bsc1�����ܴ�һ���̶��Ϸ�ӳ����ṹ��С�����ȣ� ��CHARMM�����µ�С�������ڷֲ����������ƣ� ���ֽϿ���С������[ͼ6 ��B�� ].�ɼ��� bsc1������DNAС�����ȵ�������Ϊȷ�� ��CHARMM�����²�ͬ���е�С�����Ⱦ����ھ���ṹ���ɴ˿ɼ��� AMBER���� ��bsc1�� ������DNA��С���Ŀ������Ա�CHARMM�������Ӿ�ȷ��
����2.3 �������Ӷ�DNA�ṹ��Ӱ��
����
����Ϊ��̽����ͬ�����´�С�����ȴ��ڽϴ�����ԭ�� ��DNA��Χ�������ӵķֲ�������̽����ͼ7��ͼ8�ֱ������� ��GC�� 6������ ��AT�� 6�����ڲ�ͬ������DNA�Ǽܡ���С����Χ�����ӷֲ��������ͼ7 ��A�� , ��C�� ��ͼ8 ��A�� , ��C�� �ɼ��� ������ ��GC�� 6���л��� ��AT�� 6���У� bsc1�����¹Ǽܼ�С����Χ�����ӷֲ��ܶȾ����Ը���CHARMM������bsc1�����½��ܵ����ӷֲ����õ��к�������Ǽ��ϵĸ���ɣ� ������2������Ǽ�֮��ľ����ų����ã� �����CHARMM������ȣ� bsc1�����µ�DNA�ṹС���Ŀ��Ƚ�խ���ɼ��� CHARMM��������ȷ����С�����ȵ���Ҫ�����ǣ� ��CHARMM��������������С���ķֲ����٣� ���´�����ɵ�����Ǽܾ����ų�ϴ� С�������
����
������ͼ7 ��B�� ��ͼ8 ��B�� �ɼ��� ������ͬ���У� bsc1������CHARMM������������DNA����Χ�ķֲ��ܶȲ�� Ȼ�� ��GC�� 6������ ��AT�� 6����֮��ȴ���ڽϴ���졣��������[27]������ ���� ��GC�� 6���е�DNA, �����ڴķֲ��ܶȸ��� ��AT�� 6���У� ����ģ����Ҳ�õ��˺ܺõ���֤��ͬʱ�� С����Χ�����ӷֲ����ڲ�ͬ�ļ������Ҳ���ϴ� ��ͼ7 ��C�� ��ͼ8 ��C�� �ɼ��� ���� ��AT�� 6���е�DNA, ������С���ֲ����������Ը��� ��GC�� 6���У� ������С����Χ�Ľ��ֲܷ����õ��к�������Ǽܵĸ���ɣ� �����˾����ų����ã� ʹС����խ�� ��ܺõؽ�����ʵ���� ��AT�� 6����DNA��С������С�� ��GC�� 6���е����ɼ��� ������DNA��Χ�ķֲ������Ӱ��DNA��С�����ȵ���Ҫ���ء�
����2.4ˮ���Ӷ�DNA�ṹ��Ӱ��
����
����DNA��Χˮ���ӵķֲ����ͬ����DNA�Ľṹ���нϴ��Ӱ�졣���� ��GC�� 6���У� ��ͬ������DNA��Χˮ���ӵķֲ����� ��ͼ9�� ;������ ��AT�� 6���У� CHARMM������DNA�Ĵ�С����Χ��ˮ���ӷֲ��϶� ��ͼ10�� .Ϊ��̽��ˮ���Ӷ�DNA�ṹ��Ӱ�죬 ������ȡ��ƽ��ģ����DNA�ĵ��ͽṹ�� ��ͼ3 ��B�� �ɼ��� DNAĩ�˼�������ˮ�����ƻ���Ϊ���ء���ˣ� ����CHARMM������ ��AT�� 6����ĩ�˼��������ױ���Χ��ˮ�����ƻ��� ��Ҳ�� ��AT�� 6���д�С����Χˮ���ӷֲ��϶��ԭ���ڼ��֮�����3������� ��GC�� 6���У� ������γ�2������� ��AT�� 6���и����ȶ��� CHARMM������bsc1������DNA��Χˮ���ӵķֲ������û�нϴ�����
����3������
����
�������÷��Ӷ���ѧģ���������ܼ���ķ����Ƚ���CHARMM��AMBER���� ������bsc1������OL15������ ��ˮ��Һ��B-DNA��A-DNAת�����̵�Ӱ�죬 �ֱ�õ��˲�ͬ���������ȶ���DNA�ṹ����������� ��ˮ�����в�ͬ������DNA�����ȶ��ṹ���ڲ��죬 CHARMM������DNA���ȶ��ṹ��С���Ͽ��� ����B������A����֮�䣻AMBER�����µ�DNAС����խ�� �ȶ���B���͡���һ��̽���˲�ͬ������DNA�ȶ��ṹ���ڲ����ԭ�� ���ȷ�����DNA��Χ���ӵķֲ��������������� CHARMM������DNAС����Χ�������ܶ����Ե���AMBER������ ���ܺܺõص�������Ǽܵ��ų����ã� ��CHARMM����С���Ͽ�������A���͵���Ҫԭ��һ��������ˮ���ӵķֲ���DNA�ṹ��Ӱ�죬 ����CHARMM�����µ� ��AT�� 6����ĩ�˼��֮��������ˮ�����ƻ���Ϊ���أ� ��AMBER������ȣ� CHARMM�����µ��������ý�������ģ������������ˮ��Һ��AMBER�����ܹ����õ�����ʵ�������о����Ϊ����ģ��ʵ������ṩ������ָ���� ���Խ�һ���IJ����Ż��ṩ�˰�����
����
���������
����[1]Saenger W., Defining Terms for the Nucleic Acids, Springer, New York, 1984
����[2]Ivanov V.I., Minchenkova L.E., Minyat E.E., Frank-Kamenetskii M.D., Schyolkina A.K., J.Mol.Biol., 1974, 87 ��4�� , 817-833
����[3]Mukherjee A., Lavery R., Bagchi B., Hynes J.T., J.Am.Chem.Soc., 2008, 130 ��30�� , 9747-9755
����[4]Dumont E., Wibowo M., Roca-Sanjun D., Garavelli M., Assfeld X., Monari A., J.Phys.Chem.Lett., 2015, 6 ��4�� , 576-580
����[5]Foloppe N., Mac Kerell A.D.Jr., J.Comput.Chem., 2000, 21 ��2�� , 86-104
����[6]Mac Kerell A.D.Jr., Bashford D., Bellott M., Dunbrack R.L.Jr., Evanseck J.D., Field M.J., Fischer S., Gao J., Guo H., Ha S., J.Phys.Chem.B, 1998, 102 ��18�� , 3586-3616
����[7]Ivani I., Dans P.D., Noy A., Pérez A., Faustino I., Hospital A., Walther J., Andrio P., Go1i R., Balaceanu A., Nat.Methods, 2016, 13 ��1�� , 55
����[8]ZgarbováM., Luque F.J., poner J.I., Cheatham III T.E., Otyepka M., Jurecka P., J.Chem.Theory Comput., 2013, 9 ��5�� , 2339-2354
����[9]Soares T.A., Hünenberger P.H., Kastenholz M.A., Krutler V., Lenz T., Lins R.D., Oostenbrink C., van Gunsteren W.F., J.Comput.Chem., 2005, 26 ��7�� , 725-737
����[10]Robertson M.J., Tirado-Rives J., Jorgensen W.L., J.Chem.Theory Comput., 2015, 11 ��7�� , 3499-3509
����[11]Song C., Xia Y.Y., Zhao M.W., Liu X.D., Li F., Ji Y.J., Huang B.D., Yin Y.Y., J.Mol.Model., 2006, 12 ��3�� , 249-254
����[12]Cheatham T.E., Crowley M.F., Fox T., Kollman P.A., Proc.Natl.Acad.Sci.USA, 1997, 94 ��18�� , 9626-9630
����[13]Yang L., Pettitt B.M., J.Phys.Chem., 1996, 100 ��7�� , 2564-2566
����[14]Shen W., Shao X.G., Cai W.S., Chem.J.Chinese Universities, 2016, 37 ��10�� , 1809-1816 �����ģ� ��ѧ�㣬 ���������ߵ�ѧУ��ѧѧ���� 2016, 37 ��10�� , 1809-1816��
����[15]Fu H.H., Shao X.G., Chipot C., Cai W.S., J.Chem.Theory Comput., 2016, 12 ��8�� , 3506-3513
����[16]Acosta-Reyes F.J., Subirana J.A., Pous J., Sánchez-Giraldo R., Condom N., Baldini R., Malinina L., Campos J.L., Biopolymers, 2015, 103 ��3�� , 123-133
����[17]Timsit Y., Moras D., EMBO J., 1994, 13 ��12�� , 2737
����[18]Phillips J.C., Braun R., Wang W., Gumbart J., Tajkhorshid E., Villa E., Chipot C., Skeel R.D., Kale L., Schulten K., J.Comput.Chem., 2005, 26 ��16�� , 1781-1802
����[19]Jorgensen W.L., Chandrasekhar J., Madura J.D., Impey R.W., Klein M.L., J.Chem.Phys., 1983, 79 ��2�� , 926-935
����[20]Ryckaert J.P., Ciccotti G., Berendsen H.J., J.Comput.Phys., 1977, 23 ��3�� , 327-341
����[21]Andersen H.C., J.Comput.Phys., 1983, 52 ��1�� , 24-34
����[22]Miyamoto S., Kollman P.A., J.Comput.Chem., 1992, 13 ��8�� , 952-962
����[23]Feller S.E., Zhang Y., Pastor R.W., Brooks B.R., J.Chem.Phys., 1995, 103 ��11�� , 4613-4621
����[24]Humphrey W., Dalke A., Schulten K., J.Mol.Graphics, 1996, 14 ��1�� , 33-38
����[25]Lavery R., Moakher M., Maddocks J., Petkeviciute D., Zakrzewska K., Nucleic Acids Res., 2009, 37 ��17�� , 5917-5929
����[26]Banavali N.K., Roux B., J.Am.Chem.Soc., 2005, 127 ��18�� , 6866-6876
����[27]Hamelberg D., Williams L.D., Wilson W.D., J.Am.Chem.Soc., 2001, 123 ��32�� , 7745-7755
��Ҫ��ʾ��ת�ر�վ��Ϣ��ע����Դ��985������������Ȩ�������������վ������
�Ķ���ʾ���������ж���Ϣ����ʵ�Լ��۵������վ�Ų�����
���ķ���

- �������Ŀ��ⱨ�淶�ļ���ʽҪ��
- ˶ʿ��ҵ���Ŀ��ⱨ�����Ҫ�Լ�
- �����������硷�еķ����а�����
- ��ҵ��ƿ��ⱨ�������ʽ������
- Ӣ�����Ŀ��ⱨ�淶�IJο�
- �Ŵ���ѧ���Ŀ��ⱨ�淶�IJο�
- ���̹���ѧλ���Ŀ��ⱨ����ôд
- дһƪ����ѧ˶ʿ���Ŀ��ⱨ��Ҫ
- Ӣ��˶ʿ���Ŀ��ⱨ��д������
- ������Դרҵ��ְ�о���д������
- ��ҵ�������Ʊ�ҵ�����Ŀ��ⱨ��
- ���̹���רҵ���ⱨ����ôд
- ������ְ�о������ⱨ��ע������
- һƪ3000�ֵIJ��������ְ�о���
- ��ҵ�������IJο�����д���ο���
�������
- dPCR�ķ�չ��ʷ��ԭ������Ӧ��-9
- �о���ͬ������B-DNA��A-DNA����
- ̽����ӫ��̼��Ϊӫ��Դ��������
- �����ٴ����ﻯѧ������Ŀ�в���
- HSA��֬����Ĺ�ϵ�о���T2W-RD-
- VNN1������ص�������ѧ�����о�
- PPM1A��������ø�Ľṹ�����Ե�
- DNA����ø�����ࡢ���ܼ����û�
- �㾫���Ļ�ȡ��Ӧ���������-9
- ѪС��circRNA���������о�����-
- �л�Բ���ݻ��Ե������ܻ���̽
- ���ﻯ��������״���䷢չ
- ��20����Ĥ����������о��ɹ���
- GAF�ṹ��ķ��ࡢ�ṹ������
���߿ͷ�
 ר��������ѯ
ר��������ѯ
